

Application note
Axxam S.p.A., Milan
Nanion Technologies GmbH, Munich
Intracellular ion channels are known to play an essential role in various signaling pathways for health and disease, considering that over 80% of transport processes occur inside the cells (1). Among the variety of organellar channels and transporters the proton leak channel transmembrane protein 175 (TMEM175) and the lysosomal two-pore channel (TPC) have received increasing attention in the field given their potential roles in connecting lysosomal homeostasis with pathophysiological conditions such as Parkinson’s disease and cancer (2-4). Consequently, the interest to explore intracellular ion channels as therapeutic targets has grown tremendously indicating a need for high-throughput electrophysiology including patch clamp. There has been some progress in alternative approaches such as solid supported membrane electrophysiology (SSME using the SURFE2R 96SE) recently (5), however, until now, HTS patch clamp has lacked the possibility to collect data from native lysosomes.
This is possible due to the development of special consumables (single- and multi-hole) dedicated to pursuing organellar recordings in combination with the high flexibility of the SyncroPatch 384 utilizing an ultra-low cell density approach that can use as low as 50k cells/ml, and small volumes of 1 ml for the whole of the 384-well plate, without a drastic reduction in success rate. This can be of extreme importance for expensive – as well as for samples of low quantity (cardiomyocytes, iPS cells or organelles) – to reduce costs and save time.
Our approaches resulted in the construction of cumulative concentration response curves and even intraluminal solution exchange during the recording from freshly isolated lysosomes highlighting the broad range of applications possible with the SyncroPatch 384.
Enlarged lysosomes incubated with 1 µM Vacuolin-1 were stained with 0.1 µM LysoTracker™ Red DND-99 (Invitrogen), a red fluorescent dye that stains acidic cellular compartments, such as lysosomes. The dye was added to the cells before isolation of the lysosomes and images were acquired at different magnifications and dilutions using the Operetta system (Perkin Elmer), resulting in an average diameter of 2.1 µm (Figure 1).
Figure 1
A – Isolated lysosomes stained with LysoTracker™ Red DND-99 (Invitrogen); images at different magnifications were acquired using the Operetta (Perkin Elmer).
B – Average diameter calculated at different dilutions: 3.0 ± 2.1 µm (1:10); 2.0 ± 1.3 µm (1:20); 2.4 ± 1.7 µm (1:50); data are presented as mean ± SD.
The remaining lysosomes were used on the SyncroPatch 384 for recording TMEM175 channels expressed endogenously in HEK-293 cells.
Critical for success was usage of Nanion’s “Organellar Chips”, a specialized consumable that supported and maintained the integrity of lysosomes throughout the recording and supported cumulative concentration response curves of DCPIB, a novel TMEM175 activator, able to mediate H+ and K+ currents (6) as highlighted in Figure 2. Since TMEM175 channels release luminal H+ into the cytosol, we developed assays using luminal solutions with different pH values, to enhance proton conductance, in addition to potassium flux. The seal resistance in “whole-lysosome” configuration was calculated before compound application and shows average values of 1.4 ± 0.2 GΩ and 2.1 ± 0.6 GΩ at pHluminal 4.0 and 7.0, respectively. TMEM175 activation was accompanied by a drop in Rseal, indicative for stimulation of a leak channel (Figure 2 A).
We then executed cumulative concentration additions of DCPIB to activate endogenous TMEM175 channels using only part of the NPC-384 chip (32 wells per condition). Our analysis reveals an EC50 of 65.3 ± 17.5 µM (n=5) at pHluminal 4.0 and 21.5 ± 4.1 µM (n=3) at pHluminal 7.0 for outward currents (ion and proton flux from lumen to cytosol), as shown in Figure 2 D. Representative traces (Figure 2 B-C) clearly show a larger TMEM175 current evoked in the presence of the highest DCPIB concentration in an acidic luminal environment, suggesting enhanced proton flux at acidic luminal pH.
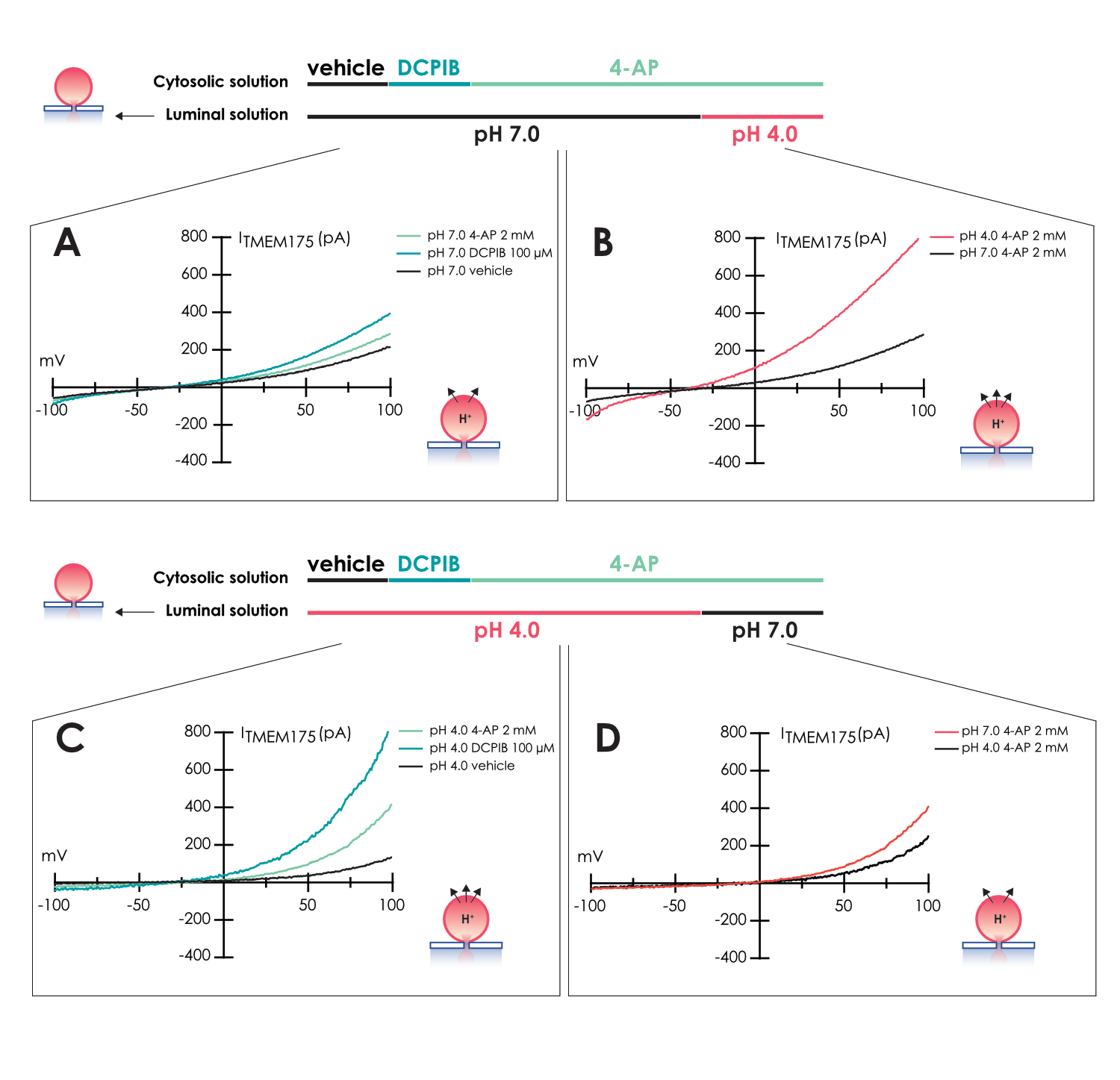
Given the known pH dependence of TMEM175 activity (7) we also employed intraluminal solution exchange for the first time where we observed a current modulation after changes in luminal pH. During the experiment with pHluminal 7.0, TMEM175 current was first evoked by DCPIB application, then partially blocked by 4-AP (Figure 3 A). In the presence of 4-AP, acidification of the luminal solution, due to the internal exchange from pHluminal 7.0 to 4.0, increases TMEM175 current (Figure 3 B).
A similar experiment was repeated by inverting the luminal pH, starting from 4.0 and changing to 7.0, using the internal perfusion feature of the SyncroPatch 384. In the presence of 4-AP, the reduction of H+ in the luminal solution induces a reduction in TMEM175 current due to a lower proton contribution (Figure 3 C-D).
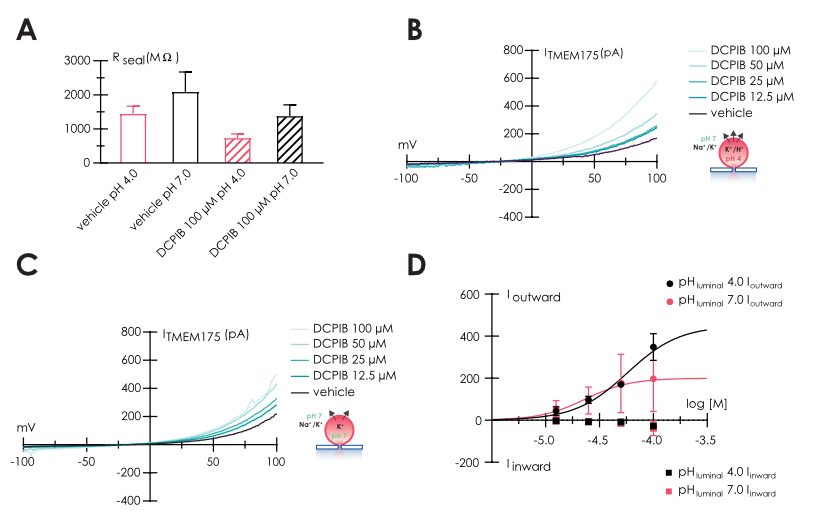
Figure 2
A – Bar graph of seal resistance calculated before and after DCPIB application.
B – Representative traces recorded in control and in the presence of increasing concentrations of DCPIB, using luminal solution with pH 4.0, and C pH 7.0.
D – Concentration response curve of DCPIB application using different luminal solution, with pH 4.0 (red) and 7.0 (black); in both experiments, cytosolic solution
was pH 7.0.
Figure 3
A – Representative TMEM175 traces recorded in control and in the presence of 100 µM DCPIB (light green) and 2 mM 4-AP (dark green); pHluminal 7.0 – pHcytosolic 7.0.
B – Effect of luminal solution exchange (from pH 7.0 to pH 4.0) on TMEM175 current in the presence of 4-AP.
C – Representative TMEM175 traces recorded in control and in the presence of 100 µM DCPIB (light green) and 2 mM 4-AP (dark green); pHluminal 4.0 – pHcytosolic 7.0.
D – Effect of luminal solution exchange (from pH 4.0 to pH 7.0) on TMEM175 current in the presence of 4-AP.
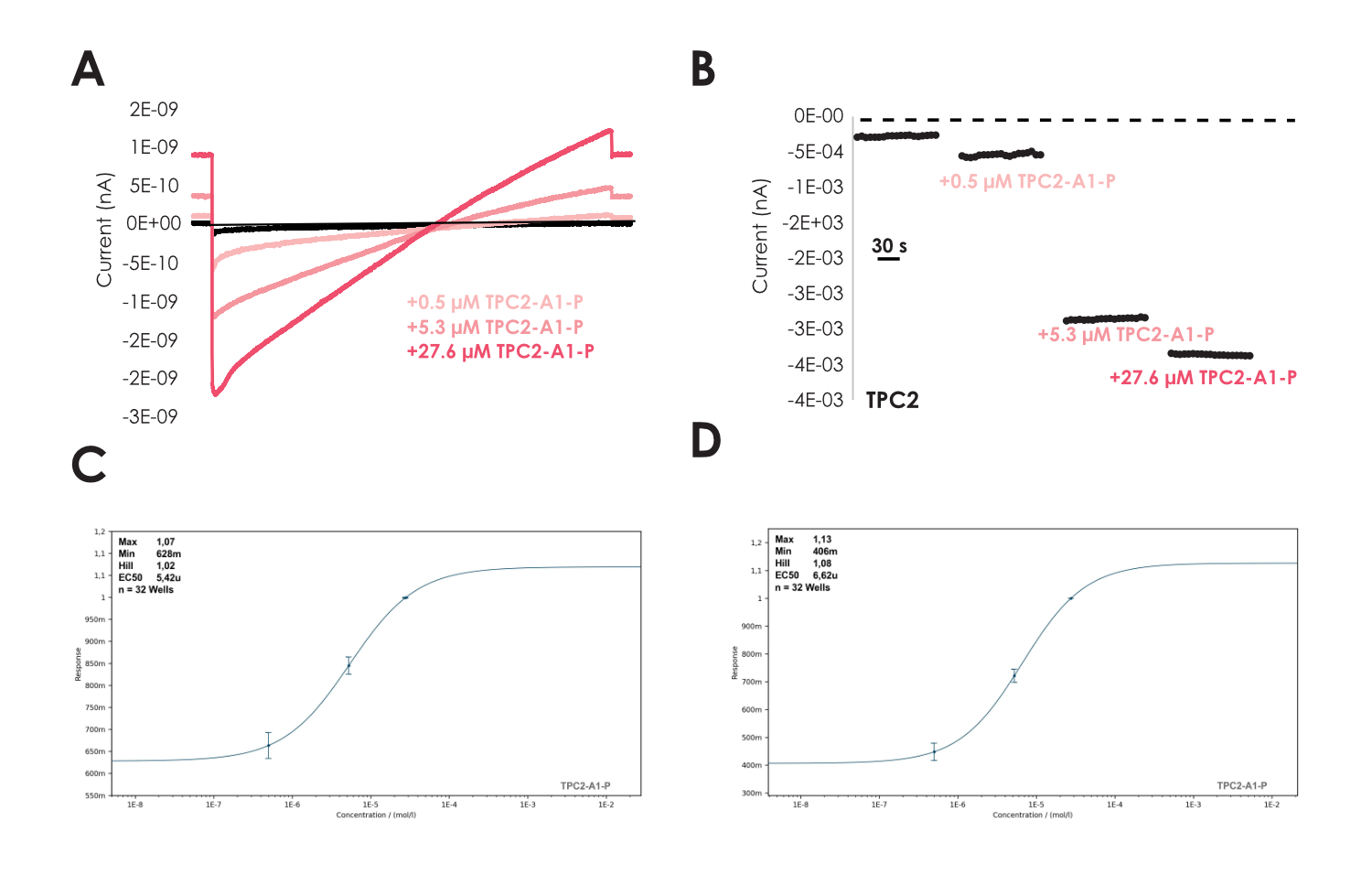
Enlarged lysosomes (Vacuolin, 1 µM) were freshly isolated as described in Schieder et al (8-9) from HEK cells either stably expressing TPC2-YFP (10-12) or without overexpression to record endogenous lysosomal currents using Nanion’s “Organellar Chips”. Lysosomes contained high amounts of TPC2 and revealed diameters in the range of few micrometers as visualized in Figure 4 A, similar to what was seen during the recordings of endogenous TMEM175 (Figure 1). Based on the epifluorescence, we could also determine a density of 100k lysosomes per ml that was sufficient to execute our studies. For TPC2, we employed the ultra-low cell density approach, where we observed lysosomal seals in ‘whole-cell’ mode that remained stable with >1 GΩ seals throughout the recording (Figure 4 B). Subsequently, we were able to elicit lysosomal currents by addition of the specific TPC2-activator TPC2_A1_P (13-14) that was absent when only reference solution was applied (Figure 4 C-D). As anticipated, the activation was accompanied by a drop in Rseal, which was not observed when reference solution was applied, indicative for stimulation of a (cation) leak channel (Figure 4 B-D). Furthermore, we performed cumulative concentration response curves on the lysosomes using TPC2_A1_P in experiments lasting ~ 15 minutes. Our analysis reveals an EC50 of 5.4 ± 1.6 µM (n = 32) for inward currents measured at -100 mV and 6.6 ± 2.7 µM (n = 32) for outward currents at +80 mV in good agreement with the literature (13-14) as highlighted in Figure 5.
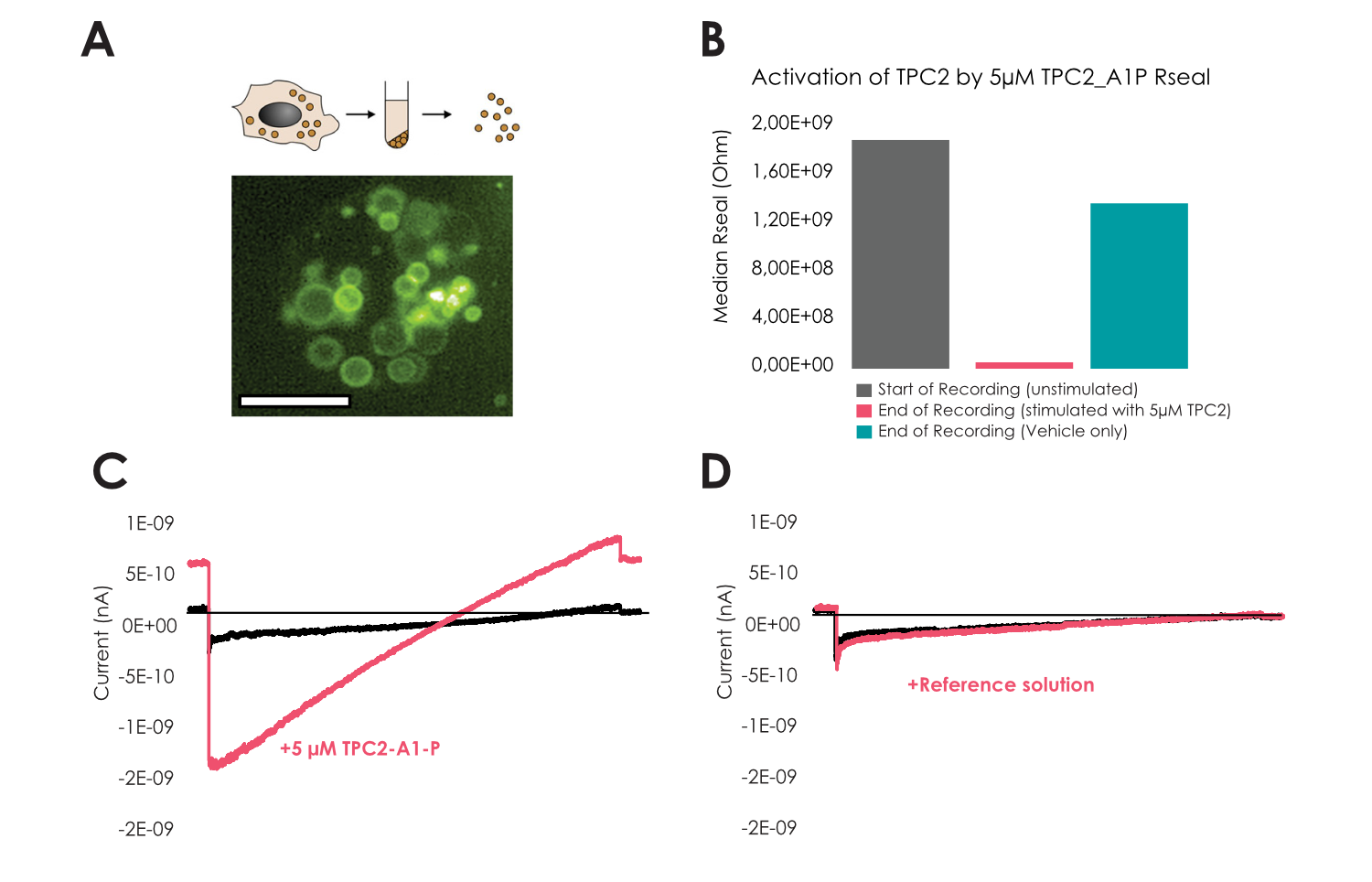
Figure 4
A – Lysosomal preparation used for the recordings from HEK cells stably overexpressing TPC2-YFP, scale bar indicates 10 µm.
B – Median Rseal at start (n=69) and end of recording when only reference solution was applied (n=15) whereas reduction in Rseal during TPC2 activation (n=54).
C-D – Exemplary current voltage ramps of Na+ currents through TPC2 in presence or absence of
5 µM TPC2_A1_P.
Figure 5
A – Example lysosomal recording applying 3 different concentrations of TPC2_A1_P as indicated.
B – Example current-time plot of inward current from recordings such as in A. Concentration response data for TPC2_A1_P at -100 mV C as well as outward current at +80 mV D yielding an EC50 of 5.4 ± 1.6 µM (n=32) and 6.6 ± 2.7 µM (n=32), respectively.
For the first time we report recordings from freshly isolated lysosomes on a high throughput automated patch clamp device, the SyncroPatch 384.
Our method satisfies the needs of primary lysosomal recordings at high throughput to explore the pharmacology of lysosomal channels in the native system. This allows a more detailed understanding of the function of lysosomal channels as well as their role as therapeutic target for numerous diseases circumventing possible caveats that stem from the detour of heterologous expression systems in the plasma membrane or usage of indirect recording approaches. We have applied our approach to record currents from lysosomes overexpressing TPC2 as well endogenous TMEM175 currents where we used specific pharmacology and tight quality control to confirm the validity of our recordings.
The results clearly indicate that the combination of our Organellar Chips/ultra low cell density method should provide efficient and low cost means of electrophysiological access to not only lysosomal channels, but also to various ion channels in other types of organelles and intracellular compartments.
It further highlights the flexibility of the SyncroPatch 384 to be adaptable and adjustable to all needs for patch clamp and allows lysosomal recording of high quality in a high throughput fashion similar to what has been observed from whole cells.
Whole cell patch clamp recordings were conducted according to Nanion’s standard procedure for the SyncroPatch 384 employing the ultra-low cell density approach and using Nanion’s Organellar Chips.
TMEM175: Traces were recorded from a holding potential of 0 mV, a voltage protocol from -100 to +100 mV with 1 s duration, was applied every 4 s to evoke TMEM175 current. Hill equation was used for EC50 calculation.
TCP2: lysosomes were held at +60mV, a voltage ramp protocol from -100 to 80 mV in 1s was applied every 10s. To estimate EC50 currents were normalized to maximum response and data fitted using the Hill equation.
Nanion Technologies GmbH
Ganghoferstraße 70a, 80339 Munich, Germany
+49 89 2190 95-0 – info@nanion.de – www.nanion.de
